
ВВЕДЕНИЕ (по книге А.Ройта)
Общие
сведения об иммунной системе
Клетки,
осуществляющие иммунный ответ
Лимфоидная система
Комплемент
Миграция
клеток и воспаление
Антитела
и клеточные рецепторы для них
Антигенраспознающие
рецепторы Т-клеток и молекулы МНС
Источники
разнообразия антигенраспознающих структур
Распознавание
антигена
Реакции клеточного
иммунитета
Взаимодействие
клеток при гуморальном иммунном ответе
Развитие иммунной
системы в онтогенезе
Регуляция иммунного
ответа
Иммунологическая
толерантность
Эволюция иммунной
системы
1 Общие сведения об иммунной системе
|
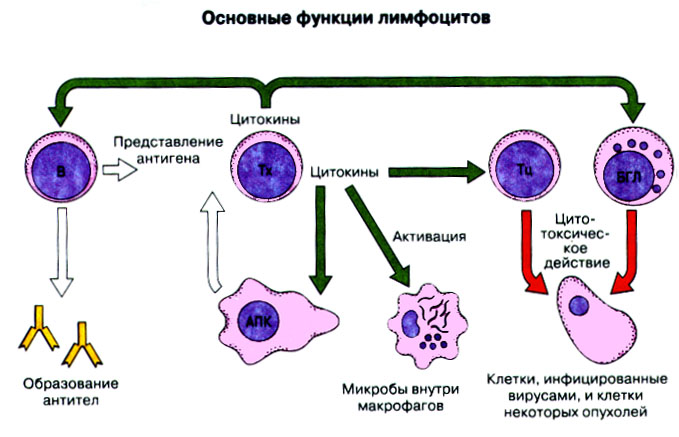
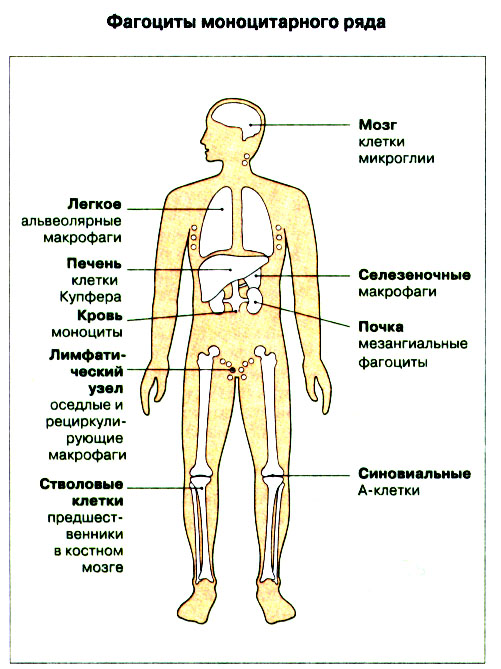
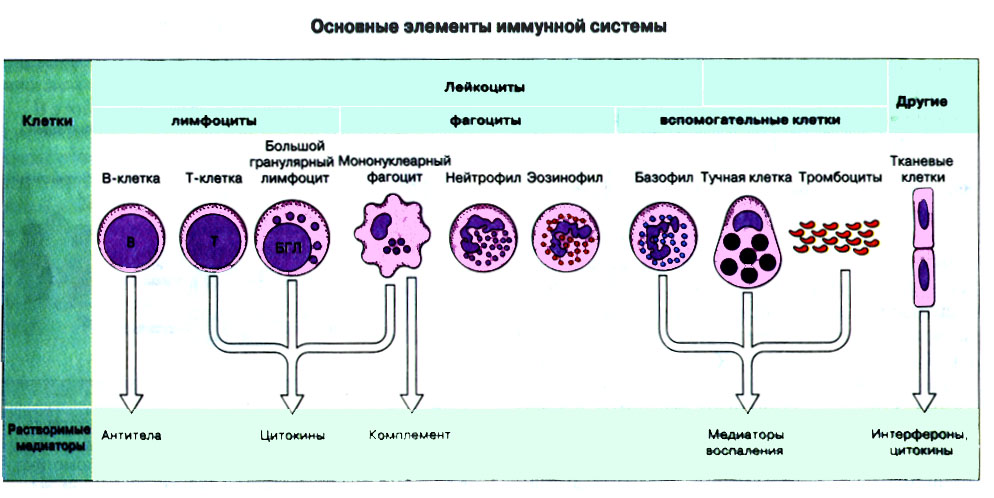
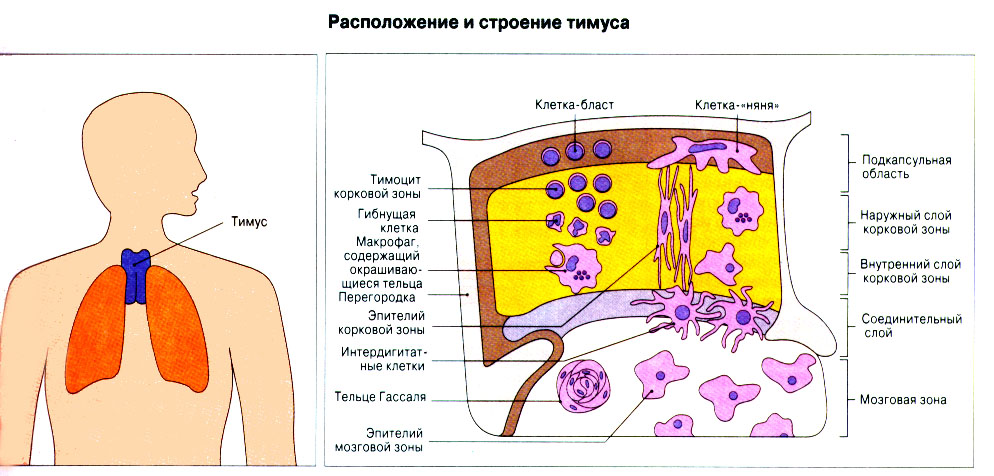
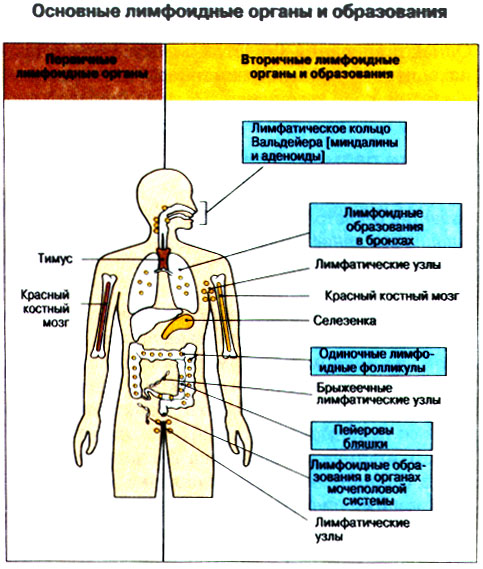
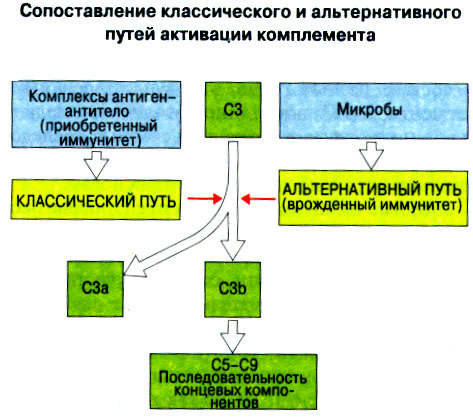
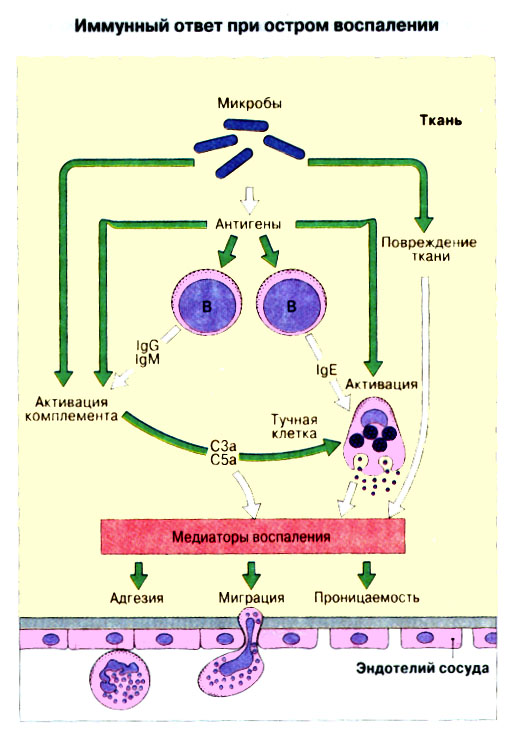
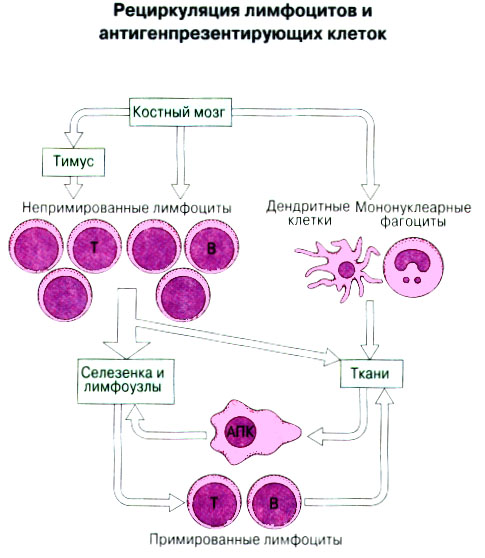
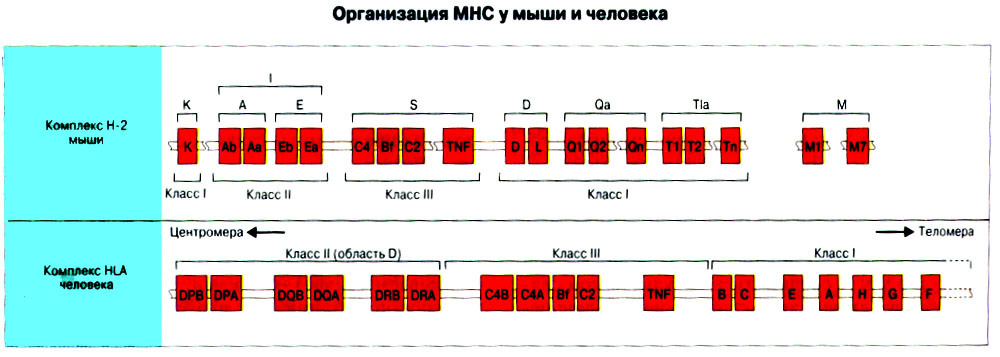
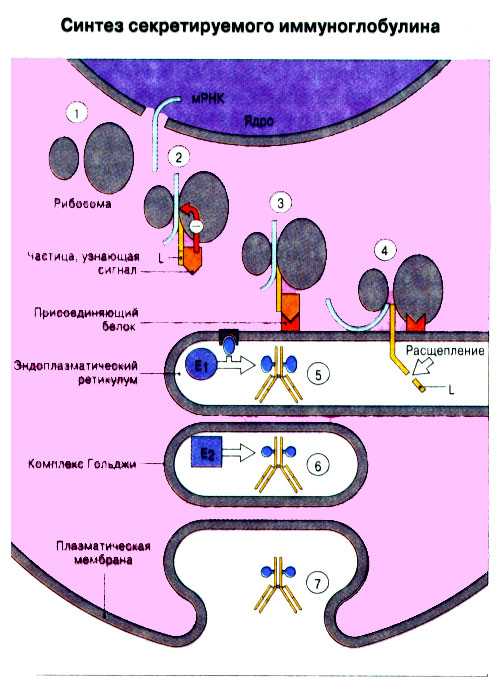
Иммунная система эволюционно формировалась для защиты макроорганизма от патогенных микробов. Одни из них, такие как вирусы, проникают внутрь клетки хозяина, другие, например многие бактерии, размножаются в тканях или полостях тела. Текст и рис. В поддержании иммунитета принимают участие лимфоциты и фагоциты. Лимфоциты распознают антигены патогенных микроорганизмов. Фагоциты поглощают и разрушают самих возбудителей. Текст и рис. Иммунный ответ состоит из двух фаз. В ранней фазе происходит распознавание антигена специфически реагирующими лимфоцитами и их активация; в поздней (эффекторной) фазе эти лимфоциты осуществляют свою координирующую функцию в устранении источника чужеродных антигенов из организма. Специфичность и память - это две основных характеристики приобретенного иммунитета. На повторную встречу с тем же антигеном иммунная система отвечает более эффективно. Лимфоциты специализированы по функциям. В-клетки образуют антитела. Цитотоксические Т-лимфоциты уничтожают клетки, инфицированные вирусами. Хелперные Т-лимфоциты координируют иммунный ответ путем контактных межклеточных взаимодействий и выделения в межклеточную среду цитокинов, которые, например, помогают В-клеткам в образовании антител. Антигены - это молекулы, распознаваемые рецепторами лимфоцитов. В-лимфоциты обычно распознают нерасщепленные молекулы антигена, тогда как Т-лимфоциты чаще всего способны распознавать антигенные молекулы только в виде фрагментов на поверхности других клеток. Узнавание молекул антигена специфичными к нему лимфоцитами влечет за собой селективное размножение лимфоцитарных клонов; клональная экспансия сопровождается дифференциацией лимфоцитов на клетки-эффекторы и клетки иммунологической памяти. При функционировании иммунной системы могут возникать нарушения, приводящие к иммунодефицитному состоянию или к гиперчувствительности, а также к аутоиммунным заболеваниям. 2 Клетки, осуществляющие иммунный ответ Многие виды клеток различного происхождения предназначены для выполнения специализированных функций в иммунном ответе. В- и Т-лимфоциты экспрессируют на своей поверхности антигенсвязывающие рецепторы и другие молекулы (маркеры), необходимые для осуществления разнообразных функций. Для Т-клеточного ответа требуется представление антигенов антигенпрезентирующими (антигенпредставляющими) клетками (АПК). Различные функциональные субпопуляции Т-лимфоцитов проявляют хелперную, супрессорную или цитотоксическую активность. РИС. Фагоцитарные клетки, несущие специфические поверхностные маркеры, циркулируют с кровотоком (моноциты и гранулоциты) и присутствуют в тканях (например, клетки Купфера в печени).РИС. Эозинофилы, базофилы, тучные клетки и тромбоциты принимают участие в воспалительной реакции. РИС. Лимфоидные органы и ткани относятся либо к первичным (центральным), либо ко вторичным (периферическим). Центральные лимфоидные органы - это тимус (РИС) и красный костный мозг(РИС). Лимфоциты дифференцируются из стволовых лимфоидных клеток в первичных лимфоидных органах и мигрируют для выполнения своих функций во вторичные лимфоидные органы и ткани. Вторичные лимфоидные органы (РИС) - это селезенка и лимфатические узлы; кроме того, в слизистых оболочках присутствуют участки вторичной лимфоидной ткани (лимфоидные образования), формирующие единую лимфоидную систему - лимфоидную ткань слизистых оболочек (ЛТС). Иммунный ответ на антигены, поступающие в организм через слизистые оболочки, начинается с примирования лимфоцитов, главным образом в пейеровых бляшках. Разные лимфоидные органы защищают различные системы организма: селезенка отвечает за антигены, циркулирующие в крови; лимфоузлы реагируют на антигены, поступающие по лимфатическим сосудам, ЛТС защищает слизистые оболочки. Лимфоциты в большинстве не оседлые, а циркулирующие клетки; они постоянно мигрируют из кровотока в лимфоидные ткани и вновь поступают в кровь через грудной лимфатический проток. Лимфоидные ткани входят в состав первичных и вторичных лимфоидных органов и образований Система комплемента - это одна из основных систем врожденного иммунитета, функция которой состоит в том, чтобы отличать "свое" от "не своего". Такая дифференциация осуществляется благодаря присутствию на собственных клетках организма регуляторных молекул, подавляющих активацию комплемента. Существует два главных пути (механизма) активации комплемента - классический и альтернативный. РИС. При классической активации происходит связывание иммунных комплексов с С1q, что соединяет приобретенный иммунитет (антитела) с врожденным (комплемент). В плазме крови постоянно происходит "холостая" активация С3, приводящая к фиксации небольшого числа его молекул на поверхности как "своего" так и "не-своего". На поверхности собственных клеток регуляторные белки вызывают разрушение связавшихся молекул С3 и подавляют дальнейшую активацию комплемента. На чужеродных структурах, лишенных регуляторных белков, напротив, начинается альтернативная активация комплемента. Наличие внутренней тиоэфирной связи в белках С3 и С4 позволяет им ковалентно взаимодействовать с гидрокси- и аминогруппами других молекул. Образование этой связи составляет ключевой момент локальной активации комплемента в очагах воспаления. При активации комплемента действуют два механизма усиления. Первый известен как "запуск ферментного каскада". "Пусковым сигналом" служит связывание небольшого числа молекул С1q, вызывающее затем последовательную активацию ряда зимогенов (проферментов), которые расщепляют уже значительно большее число молекул С3. Второй механизм усиления - это действующая по принципу положительной обратной связи "петля усиления". Расщепление небольшого количества молекул С3 с образованием С3b способствует появлению фермента С3-конвертазы, который расщепляет гораздо больше С3. На собственных клетках организма имеются молекулы, подавляющие действие этой петли усиления путем расщепления С3b на неактивные продукты. На чужеродных структурах действие "петли усиления" не встречает препятствий. Эффекторные механизмы системы комплемента делятся на пять групп в зависимости от функции:
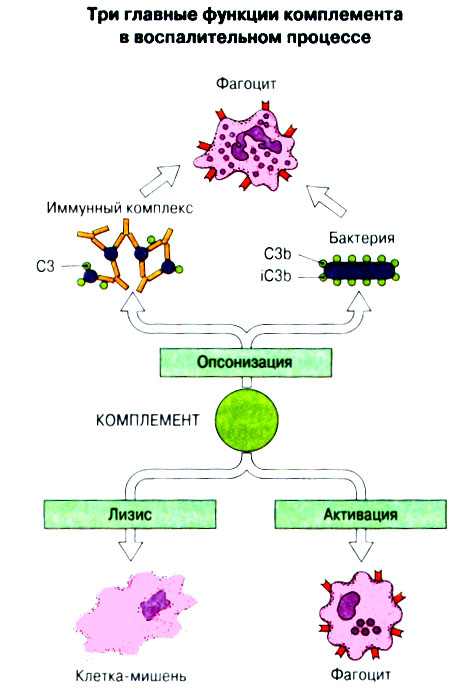
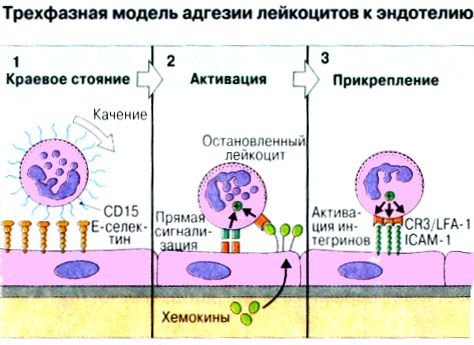
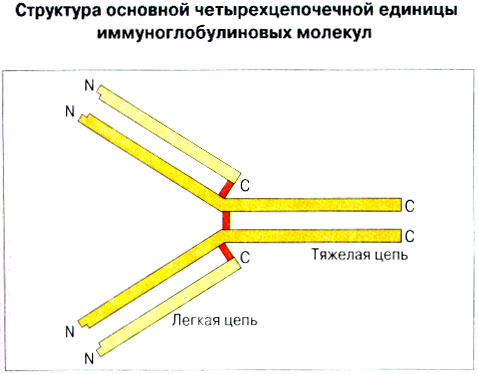
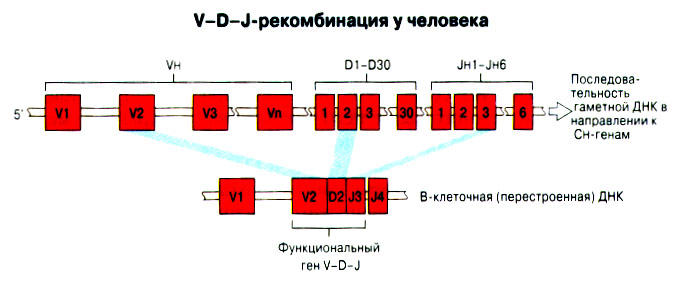
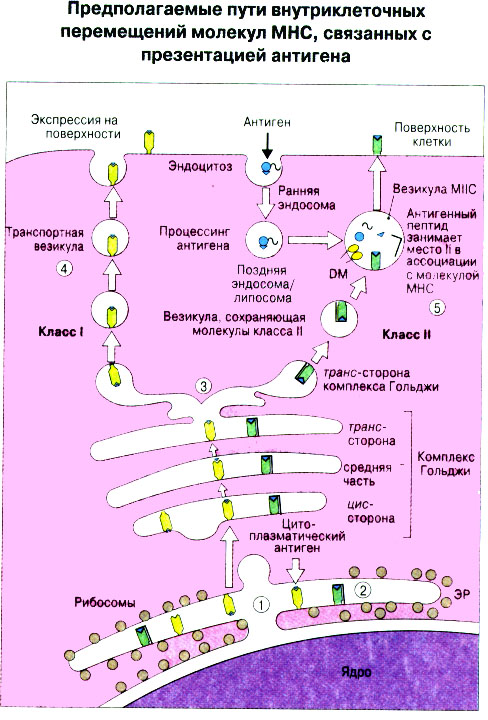
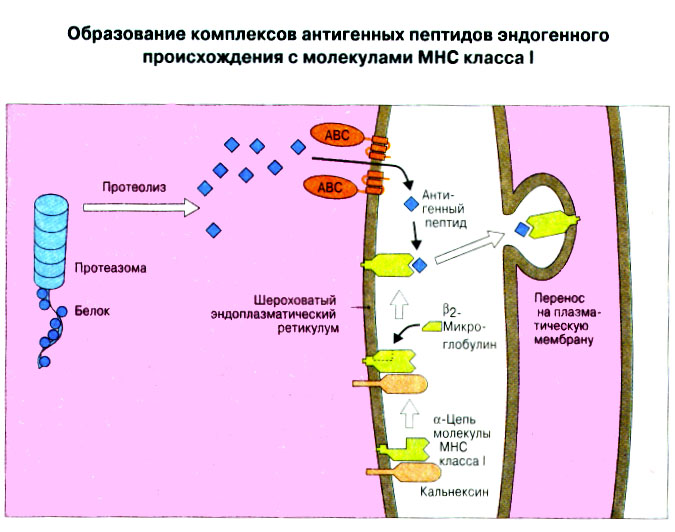
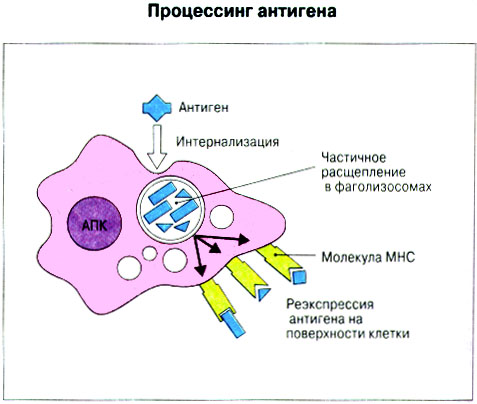
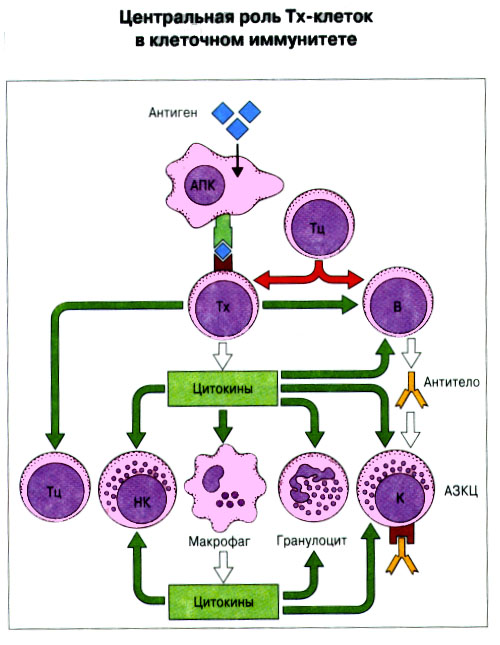
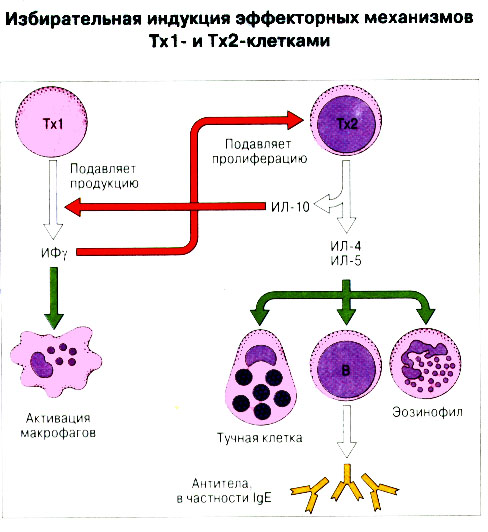
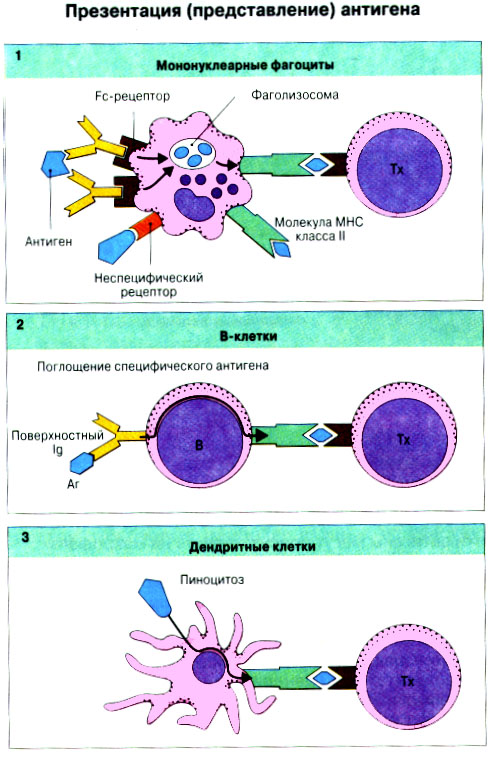
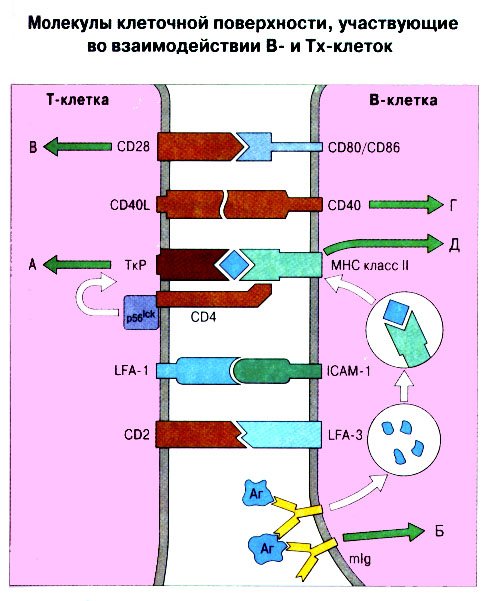
Патогенные микробы вырабатывают механизмы, позволяющие им избежать уничтожения системой комплемента, а в некоторых случаях даже использовать для усиления своей патогенности. Система комплемента может участвовать в патогенезе заболевания, если происходит ее генерализованная активация in vivo или активация на собственных тканях в результате связывания комплемента аутоантителами. Опсонизация. Это стимуляция фагоцитоза в результате прикрепления белков комплемента к поверхности мишеней (микробов, иммунных комплексов и др.). Обладая рецепторами к опсонизирующим белкам, фагоцитарные клетки связывают мишени, что вызывает активацию фагоцитов и эндоцитоз или фагоцитоз мишеней. Хемотаксис (направленное движение клеток). Активация лейкоцитов. Полиморфноядерные гранулоциты и макрофаги обладают специфическими рецепторами к мелким фрагментам белков комплемента, образующимся на поверхности мишеней в результате каскада протеолитических реакций. Диффундируя в окружающую среду, эти фрагменты привлекают фагоциты и, связываясь с ними, вызывают их активацию.Рис.5.12. Лизис клеток-мишеней. Протеолитический каскад комплемента завершается погружением гидрофобного "зонда" в липидный бислой мембраны клетки-мишени и ее последующим осмотическим разрывом и лизисом. РИС. 5 Миграция клеток и воспаление Воспаление - это реакция организма, обеспечивающая привлечение лейкоцитов и растворимых компонентов плазмы в очаги инфекции или повреждения тканей. К его основным проявлениям относятся повышение кровенаполнения капилляров и их проницаемости для сывороточных макромолекул, а также усиленная миграция лейкоцитов в направлении воспалительного очага через эндотелий расположенных поблизости сосудов.РИС. Миграция лейкоцитов представляет собой сложный процесс, который зависит от особенностей данной популяции мигрирующих клеток, уровня активации клеток и характера их взаимодействия с эндотелием в разных участках сосудистого русла.РИС. Путь миграции частично определяется уровнем активации клеток: покоящиеся или еще не встречавшие антиген лимфоциты имеют тенденцию мигрировать через венулы с высоким эндотелием в лимфоидные ткани, тогда как активированные лимфоциты - в очаги воспаления. Молекулы адгезии, регулирующие миграцию лейкоцитов, по структурным признакам относятся к разным, но родственным семействам, в том числе к суперсемейству иммуноглобулинов (эндотелиальные молекулы клеточной адгезии), семейству или семейству интегринов. В эндотелиальных клетках синтез молекул адгезии индуцируют цитокины. Экспрессия лейкоцитарных молекул адгезии зависит от популяции клеток и стадии их дифференцировки. РИС Хемотаксические молекулы определяют направление миграции лейкоцитов и, кроме того, инициируют этот процесс при контакте лейкоцитов с поверхностью эндотелия. Медиаторы воспаления, выделяемые тучными клетками, тромбоцитами и лейкоцитами при иммунологических реакциях организма или повреждении тканей, действуют совместно с продуктами активации ферментных систем плазмы, осуществляя регуляцию проницаемости и кровенаполнения сосудов. 6 Антитела и клеточные рецепторы для них Циркулирующие антитела распознают антиген в крови и в тканевой жидкости. У большинства видов млекопитающих пять классов антител - IgA, IgG, IgM, IgD и IgE. Основная структурная единица иммуноглобулинов состоит из двух легких и двух тяжелых цепей. Классы различаются между собой тяжелыми цепями. IgA и IgM - это олигомеры основной четырехцепочечной единицы. РИС Цепи иммуноглобулинов свернуты в несколько глобулярных структур, называемых доменами; легкие цепи образуют по два домена, тяжелые - четыре или пять в зависимости от класса Ig. С помощью протеолитических ферментов можно получать фрагменты иммуноглобулинов для исследовательских либо медицинских целей. Папаин расщепляет молекулу IgG на три фрагмента - два антигенсвязыващих (Fab) и один Fc; пепсин отепляет крупный F(ab')2 - фрагмент, содержащий оба антигенсвязывающих центра. Антигенсвязывающие центры образованы гипервариабельными (V) участками цепей Ig. В V-доменах любой легкой или тяжелой цепи имеется по три таких участка. Свертывание цепей в домены приводит к тому, что гипервариабельные участки группируются на выступающих частях молекулы, образуя два антигенсвязывающих центра в каждой четырехцепочечной единице. Все антитела несут две функции. Кроме связывания антигена, они осуществляют одну или несколько эффекторных функций. Структурные участки молекулы иммуноглобулина, ответственные за эффекторную активность (например, за активацию комплемента или связывание с клеткам), пространственно удалены от антигенсвязывающих центров и находятся главным образом в Fc-области. Рецепторы для иммуноглобулинов присутствуют на поверхности мононуклеарных лейкоцитов, нейтрофилов, нормальных клеток-киллеров, эозинофилов, базофилов и тучных клеток. Взаимодействуя с Fc-областью иммуноглобулинов разных изотипов, рецепторы стимулируют, например, фагоцитоз, противоопухолевую цитотоксическую активность и дегрануляцию тучных клеток. Большинство Fc гамма-рецепторов относится к молекулам иммуноглобулинового суперсемейства и имеет два или три внеклеточных иммуноглобулин-подобных домена. 7. Антигенраспознающие рецепторы Т-клеток и молекулы МНС Антигенраспознающий Т-клеточный рецептор - это гетеродимерный (альфа-бета или гамма-дельта) гликопротеин, благодаря которому Т-клетки способны распознавать разнообразные антигены. На поверхности Т-клеток он ассоциирован с полипептидным комплексом, имеющим групповое название CD3. Главный комплекс гистосовместимости (MHC) РИС кодирует два набора высокополиморфных белков клеточной поверхности, названных молекулами MHC класса I и класса II. Альфа-бета ТкР распознает процессированный антиген в виде пептидных фрагментов, связанных с молекулами MHC класса I или II. При этом и молекула MHC, и фрагмент антигена контактируют с ТкР. Укладка полипептидных цепей молекулы MHC обеспечивает образование полости, в которой связываются пептидные фрагменты процессированного антигена для их презентации Т-клеткам. Молекулы класса I способны связывать пептиды из 8-9 аминокислотных остатков, молекулы класса II - несколько более длинные. Карманы антигенсвязывающей полости могут вмещать пептиды различной структуры в зависимости от гаплотипа MHC. Высокий полиморфизм молекул MHC, а также способность каждой АПК экспрессировать несколько разных молекул MHC обеспечивают возможность презентации Т-клеткам множества самых различных антигенных пептидов. 8. Источники разнообразия антигенраспознающих структур Благодаря огромному разнообразию антител, синтезируемых В-клетками, и антигенраспознающих рецепторов, экспрессируемых Т-клетками, иммунная система способна распознавать множество различных антигенов и отвечать на них. Молекула иммуноглобулина состоит из тяжелых и легких цепей; легкие цепи могут относиться к каппа- или лямбда-типу. Общее количество возможных вариантов антигенсвязывающих центров рассчитывается как произведение чисел различных тяжелых и легких цепей. Легкие цепи иммуноглобулинов кодируются генными сегментами V и J; в кодировании тяжелых цепей участвуют сегменты V и J, но дополнительное разнообразие вносят сегменты D. РИС Рекомбинации ограниченного числа генных сегментов V, D и J создают бесконечное число вариабельных доменов разной специфичности. После антигенной стимуляции в генах легких и тяжелых цепей иммуноглобулинов происходят точечные соматические мутации. Гены Т-клеточных рецепторов при этом не подвергается изменениям. В кодировании ТкР участвуют четыре группы генов: гены альфа и бета экспрессирует большинство периферических Т-клеток, а гены гамма и дельта - одна из субпопуляций Т-клеток тимуса и небольшая часть периферических Т-клеток. Разнообразие ТкР, подобно разнообразию антител, создается в результате рекомбинаций между генными сегментами V, D и J, происходящих в каждом из локусов альфа-, бета-, гамма- или дельта-цепей с небольшими различиями в механизмах. Рекомбинацию генных сегментов V, D и J, кодирующих иммуноглобулины и Т-клеточные рецепторы, регулируют (по крайней мере отчасти) два активирующих ее гена (RAG-1 и RAG-2). Кроме простых перестановок генных сегментов V, D и J в создании разнообразия иммуноглобулинов и ТкР имеют значение вставки добавочных нуклеотидов ("N-региональная" вариабельность), изменение позиций стыковки генных сегментов и рамок считывания сегментов D. Переключение изотипов иммуноглобулинов обусловлено рекомбинацией VDJ-генов с различными С-генами и дифференциальным сплайсингом РНК. РИС Антитела высокоспецифичны по отношению к трехмерной конформации эпитопов антигена, вызвавшего их образование. Аффинность (сродство) антитела - мера прочности связи антигенсвязывающего центра молекулы антитела с отдельным эпитопом антигена. Функциональная аффинность, или авидность, взаимодействия антител с антигеном определяется также числом антигенсвязывающих центров в молекуле антитела и их способностью связываться с многочисленными эпитопами данного антигена. Т-клетки распознают связанные (презентируемые) другими клетками антигены в ассоциации с молекулами MHC класса I или класса II. РИС Пептидные фрагменты процессированных антигенов связываются в специальной полости молекул MHC. Молекулы MHC класса I или класса II презентируют пептиды соответственно эндогенных и экзогенных антигенов. В зависимости от своего происхождения процессированные антигены встречаются и связываются с молекулами MHC в различных внутриклеточных органеллах. Антигенные пептиды, связываемые молекулами MHC класса I, образуются в цитоплазме в результате расщепления антигенов органеллами, названными протеазомами. Движение этих пептидов по эндоплазматическому ретикулуму обеспечивают транспортные белки из суперсемейства "АВС". Комплекс из трех компонентов - тяжелая цепь класса I - бета 2-микроглобулин - пептид - перемещается на клеточную поверхность. РИС Антигенные пептиды, связываемые молекулами MHC класса II, образуются из поглощенных путем эндоцитоза экзогенных антигенов, в результате их процессинга в эндосомах или лизосомах. Молекулы MHC класса II в составе комплекса с инвариантной (Ii) полипептидной цепью транспортируются через комплекс Гольджи в эндосомы, где утрачивают Ii-цепь в результате диссоциации и присоединяют антигенные пептиды. РИС Комплексы антигенных пептидов с молекулами MHC, экспонированные на клеточной поверхности, могут распознаваться специфическими рецепторами Т-клеток. Однако для последующей активации Т-клеток требуется ряд дополнительных взаимодействий с участием вспомогательных молекул. 10. Реакции клеточного иммунитета Цитокинам принадлежит центральная роль в положительной и отрицательной регуляции иммунного ответа, а также в его интеграции с физиологическими функциями других систем организма - эндокринной и гемопоэтической. Распознавание микробных структур происходит в самом начале реакции организма на инфекцию, до развития специфического иммунного ответа. Тип последующего ответа зависит в основном от выделяемых цитокинов. Регуляцию иммунного ответа осуществляют хелперные Т-клетки (Тх). РИС Отвечая на антиген, они выделяют различные наборы цитокинов и тем самым инициируют разные эффекторные функции. Так, Тх1-клетки активируют макрофаги, а Тх2-клетки способствуют образованию антител. Если активирована неадекватная эффекторная функция, элиминации возбудителя не происходит и развивается хроническая иммунопатология. Иммунный ответ Тх1-типа подавляет ответ Тх2-типа, и наоборот. РИС Большинство цитотоксических Т-клеток распознает антиген, презентированный в ассоциации с молекулами MHC класса I, тогда как НК-клетки реагируют на мишени, не экспрессирующие эти молекулы. Цитотоксическая активность клеток-киллеров - это комбинированное воздействие на клетки-мишени путем прямого контакта, выделения цитокинов и экзоцитоза белков из гранул, в частности перфорина и гранзимов. Активированные макрофаги уничтожают поглощенные ими микроорганизмы с помощью высокоактивных метаболитов кислорода и азота. Когда реакции клеточного иммунитета не обеспечивают устранения инфекции или персистирующего антигена и поэтому не могут завершиться, в тканях возникает хронический деструктивный воспалительный процесс или образуются гранулемы. При этом непосредственное разрушение жизненно важных клеток или вторичные микрососудистые нарушения, обусловленные избыточным выделением цитокинов, могут стать причиной иммунопатологии. 11. Взаимодействие клеток при гуморальном иммунном ответе Иммуноактивация, необходимая для синтеза антител, включает взаимодействие между Т-клетками и АПК и затем между этими примированными Т-клетками и В-клетками. РИС Активация клеток осуществляется путем антигенспецифичного взаимодействия с участием молекул клеточной адгезии и цитокинов. Наиболее сильным костимулирующим сигналом служит молекула В7 (CD80 или CD86). РИС В отсутствие соответствующей костимуляции распознавание антигена может привести к развитию клональной анергии. Пролиферация лимфоцитов происходит опосредованно, т.е. зависит от индукции рецепторов для факторов роста лимфоцитов, которую вызывает активация. Факторы роста лимфоцитов (например ИЛ-2) синтезируются главным образом Т-клетками. Существуют два типа антигенов, вызывающих гуморальный иммунный ответ - Т-зависимые и Т-независимые. Т-зависимые антигены индуцируют вторичный иммунный ответ, характеризующийся образованием IgG и повышением аффинности антител. Для первичного иммунного ответа на Т-зависимые антигены характерно образование низкоаффинных IgM-антител. При вторичном иммунном ответе продуцируется большее количество антител и происходит переключение изотипов с образованием IgG, IgA и IgE, Одновременно с этим возрастает аффинность антител. Переключение изотипа и повышение (созревание) аффинности антител происходит в центрах размножения внутри вторичных лимфоидных тканей. Гуморальный иммунный ответ (образование антител) представляет собой кульминацию ряда клеточных и молекулярных взаимодействий, происходящих в определенной последовательности:
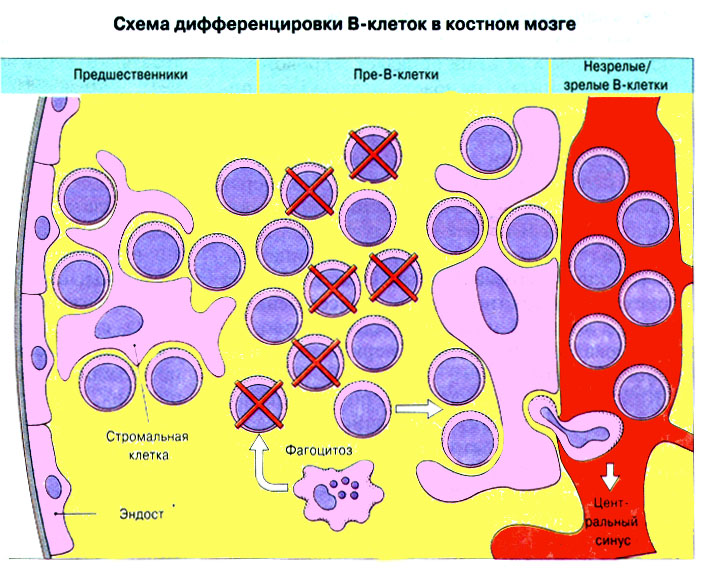
12. Развитие иммунной системы в онтогенезе Большинство клеток иммунной системы образуется из гемопоэтических стволовых клеток. РИС Развитие клеток разных типов (рядов, или линий, дифференцировки) зависит от межклеточных взаимодействий и цитокинов. Лимфоидные стволовые клетки развиваются и созревают в первичных лимфоидных органах. Этот процесс называется лимфопоэзом. Т-лимфоциты при развитии в тимусе подвергаются положительной и отрицательной селекции. В-клетки млекопитающих развиваются главных образом в печени плода, а после рождения - в костном мозге. Этот процесс прдолжается в течение всей жизни организма. В-клетки в местах своего развития также подвергаются отбору. Репертуар антител, существующий у взрослых особей, формируется в процессе лимфопоэза путем рекомбинации генных сегментов, кодирующих ТкР и Ig. В центрах размножения происходит олигоклональная пролиферация В-клеток, переключение изотипа антител, повышение аффинности и формирование иммунологической памяти. 13. Регуляция иммунного ответа Иммунный ответ регулируется разнообразными механизмами, которые обеспечивают восстановление исходного состояния иммунной системы, после того как реакция на данный антиген перестает быть необходимой. Конечный результат любого иммунного ответа зависит от многих факторов, в том числе от свойств антигена, его дозы и пути поступления, а также от генетических особенностей организма. Иммуноглобулины могут играть в иммунном ответе положительную роль, действуя как антиидиотипические антитела или образуя иммунные комплексы. Возможна и отрицательная роль Ig в иммунном ответе, когда они ослабляют антигенный стимул, маскируя детерминанты антигена в результате связывания или способствуя выведению антигена из организма. Иммунный ответ может зависеть от способности антигенпрезентирующих клеток обеспечивать костимуляцию Т-лимфоцитов. Регуляцию иммунного ответа способны осуществлять Т-клетки. Опыты с переносом иммунокомпетентных клеток показали, что Т-лимфоциты CD4+ могут подавлять последующие иммунные ответы. Регуляторный эффект оказывают и Т-клетки CD8+. Продукция цитокинов Т-лимфоцитами влияет на тип иммунного ответа, вызываемого антигеном. Генетически иммунный ответ зависит как от генов MHC, так и от не относящихся к MHC генов. Кроме того, поскольку на него влияет нейроэндокринная система, он зависит и от генетических факторов, определяющих функции этой системы. РИС Эффективный иммунный ответ - результат взаимодействия между антигеном и целой сетью иммунокомпетентных клеток. Факторы, влияющие на иммунный ответ:
14. Иммунологическая толерантность Иммунологическая толерантность - это состояние ареактивности в отношении того или иного антигена; ее индуцирует предшествующий контакт с этим антигеном. Свойство различать "свое - несвое" появляется в онтогенезе; все эпитопы (антигенные детерминанты), закодированные в ДНК организма, должны быть иммунологически определены как "свои", все другие - как "несвои". Механизмы толерантности необходимы, поскольку иммунная система продуцирует огромное число разнообразных антигенспецифичных рецепторов и некоторые из них оказываются специфичными к собственным антигенам организма; толерантность предотвращает нежелательные реакции против собственных органов и тканей. Центральная (тимическая) толерантность к своим антигенам (аутоантигенам) обеспечивается делецией тех дифференцирующихся Т-клеток, антигенспецифичные рецепторы которых обладают высоким сродством к собственным антигенам, локализованным в тимусе. Низкоаффинные аутореактивные Т-клетки, а также Т-клетки с рецепторами к тем антигенам, которые не представлены в тимусе, созревают и пополняют пул периферических Т-лимфоцитов. Посттимическую толерантность к собственным антигенам обеспечивают три механизма:
Делеция В-клеток происходит в костном мозге; делетируются на ранней стадии дифференцировки те В-клетки, которые экспрессируют на своей поверхности иммуноглобулиновые рецепторы с высокой аффинностью к собственным мембраносвязанным антигенам. Аутореактивные В- и Т-клетки могут избежать делеции на периферии за счет снижения экспрессии антигенных рецепторов. Толерантность можно индуцировать искусственно различными способами, и некоторые из них применимы в медицине для предотвращения отторжения чужеродных трансплантатов и лечения аутоиммунных и аллергических заболеваний. Кроме структурных особенностей эпитопов важное значение при определении "свое - не свое" имеют следующие факторы:
У беспозвоночных важную роль в ликвидации чужеродного материала выполняют фагоцитоз и инкапсулирование; у многих видов эти процессы осуществляются белыми клетками крови. Установлена возможность распознавания чужеродных трансплантатов у животных, появившихся уже на ранних этапах эволюции, но специфичность реакции и клональная экспансия клеток у беспозвоночных недостаточно доказаны. В процессах распознавания "свое/не-свое" участвуют лектины и профенолоксидаза. У многих беспозвоночных в иммунорегуляции участвуют цитокин-подобные молекулы. Иммуноглобулинами беспозвоночные не обладают , хотя у многих из них обнаружены IgG-подобные молекулы. У некоторых вторичнополостных наблюдаются индуцируемые гуморальные иммунные реакции широкого спектра. У всех челюстноротых позвоночных имеются В-клетки и IgM. У пойкилотермных животных, несмотря на присутствие дополнительных тяжелых цепей, не относящихся к мю-цепям, афффинность антител остается низкой. Метод рекомбинантной ДНК позволяет обнаружить у позвоночных разных уровней филогенеза несколько типов организации генов Ig. У хрящевых рыб идентифицирован главный комплекс гистосовместимости (MHC). Начиная с костных рыб, появляются Т-клетки; рецепторные молекулы этих клеток в настоящее время изучаются. К элементам системы врожденного иммунитета у позвоночных относятся нормальные клетки-киллеры (НК), фагоциты, белки комплемента и иммунорегуляторные цитокины.
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}